Jerry Blythe1 and Marco Ruggiero2
1Retired Medical Doctor, Indianapolis, IN, USA
2National Coalition of Independent Scholars, San Antonio, TX, USA
Corresponding author Marco Ruggiero, MD, PhD. National Coalition of Independent Scholars, San Antonio, TX, USA
E-mail: marco.ruggiero@ncis.org
Abstract
In this study, we describe the changes associated with the consumption of an extremely biodiverse probiotic yogurt in a 55-year-old female from South Vietnam. In August 2019, the subject voluntarily embarked on a three-month nutritional experience and decided to share her experience with the goal of advancing scientific knowledge in the field of nutritional health. Consumption of this biodiverse probiotic yogurt was associated with a decrease in serum alpha- N-acetylgalactosaminidase (nagalase) activity, increased elimination of toxic metals and non- metal toxicants, a trend toward normalization of the lipid profile, and a trend toward a rebalance of the gut microbiota.
Key words: probiotic yogurt; nagalase; detoxification; toxic metals; microbiome
Introduction
We have previously observed and reported in two recent articles (1,2) that the consumption of an extremely biodiverse probiotic yogurt with a unique microbial composition is associated with detoxification of non-metal toxicants (3), decrease of serum alpha-N- acetylgalactosaminidase (nagalase) activity (3,4), decrease of serum C-reactive protein (CRP) (4), decrease of markers specific for multiple myeloma (5), and a dramatic decrease of viral load in Hepatitis B (6).
In this report, we describe the changes associated with consumption of an extremely biodiverse probiotic yogurt in a 55-year-old female from South Vietnam who presented in August 2019 feeling sometimes feverish at night with morning fatigue for several months.
Subject Information
The subject grew up in South Vietnam during the Agent Orange era and left Vietnam at age 17 as part of the “Boat Migration” in the 1970s. Settling in the United States, she earned a college degree in mathematics and computer sciences, and has worked in the Chicago area the past 35 years. The subject is a Buddhist minister, active in her Temple with several outside business interests. She described hour-long commutes for years to and from downtown Chicago to her alternative medical clinic.
She is not a runner but ran the 2019 Peachtree 10 Km road race in Atlanta on July 4 in 90 degree fahrenheit weather without incident. She is married and raised three children. As a teenager she suffered from severe nutritional anemia with delayed menses until 17 (hemoglobin reported 6 g/dL requiring hospitalization and units of blood). The subject returned to South Vietnam in 2016 and again in 2018. Upon returning from her last trip, she described vague abdominal symptoms and not feeling well.
She reported mold exposure, a common finding living in the midwestern part of the United States. She also reported a persistent mild dry cough, sneezing, and sinus congestion, along with fatigue and a poor sleep cycle.
In 2017 the subject had the flu and was bedridden for 6 weeks. It is worth mentioning that 2017 was a record year for the flu in the United States with a significant number of deaths (7). The subject reported few other lifetime illnesses or problems. In August 2019, the subject voluntarily embarked on a three-month nutritional experience that included consumption of an extremely biodiverse probiotic yogurt, as described in references 3-6.
Study Design and Methodology
The subject provided one of the authors Dr. JB selected laboratory studies from 2016 through 2019 to go along with her current studies with the goal of advancing scientific knowledge in the field of nutritional approaches to health. In addition to using the biodiverse probiotic yogurt
Blythe and Ruggiero 2020 – Probiotics and detoxification
2
during her three-month experience, the subject reported taking several supplements including a multi-vitamin, 2000 to 5000 IU vitamin D3 daily, Omega-3, glucosamine and recently a non- prescription thyroid supplement containing vitamins and minerals to assist her morning fatigue. It is worth noting that the supplements were consumed both before and during the experience with the regular consumption of the probiotic yogurt. It is therefore plausible that the observed changes are attributed mainly to the consumption of the probiotic yogurt.
Blood and urine analyses were performed at different times before this study and after the three-month experience. Determination of serum alpha-N-acetylgalactosaminidase (nagalase) activity was performed at the Health Diagnostics and Research Institute (South Amboy, NJ, USA), a consociate of European Laboratory of Nutrients, (Bunnik, The Netherlands); the results are expressed as nMol/mL/min. Blood analyses were performed at Lab Corp (Laboratory Corporation of America) and Quest Diagnostics. Analyses of metal and non-metal toxicants in early morning urine samples were performed at The Great Plains Laboratory Inc. (Lenexa, KS, USA) and were normalized per grams of creatinine. Copies of the original records are conserved at the office of Dr. JB. In this article we describe and discuss only the values that changed during the three-month experience and, for the sake of brevity, we omit discussing the values that did not change.
Consent
Since this is a single case report that does not produce generalizable knowledge, nor is it an investigation of an FDA regulated product, it is accepted that Institutional Review Board (IRB) review is not required for this activity (see, for example, ref. 8). Written informed consent for publication of clinical details and laboratory data was obtained from the subject and it is conserved at the office of Dr. JB.
Results
Serum lipid profile
Comprehensive Metabolic Profile Studies by Lab Corp were performed prior to this experience in 2018 and 2019. A lipid profile in July 2018 showed high triglycerides (375 mg/dL; reference range: 30-150 mg/dL), mildly elevated total cholesterol (231 mg/dL: reference range: <199 mg/dL), and normal blood sugar. Triglyceride values improved to 232 mg/dL over the three- month experience with the probiotic yogurt, and cholesterol fell to 209 mg/dL, only slightly elevated. These results seem to indicate that consumption of the probiotic yogurt was associated with a trend toward normalization of the lipid profile and are consistent with previous observation concerning probiotic yogurts and serum lipids (9).
Serum vitamin D3
Vitamin D3 was quite low in 2017 at 15 ng/ml (reference range: 30-100 ng/ml). Levels improved in 2018 to 37 ng/ml and to 43.5 ng/ml in August 2019, and to 100 ng/ml in December 2019
Blythe and Ruggiero 2020 – Probiotics and detoxification
3
following the three-month experience with the biodiverse probiotic yogurt. It is worth mentioning, however, that, since beginning the experience in 2019, the subject began walking outdoors in the sun 30 to 45 min two to three days per week and consumed a 2000 to 5000 IU vitamin D3 supplement. It is plausible, therefore, that the normalization of vitamin D3 values has to be ascribed primarily to consumption of the vitamin supplement and exposure to sunlight with possible contribution from the probiotic yogurt (10).
Serum nagalase activity
Serum nagalase activity was elevated prior to the three-month experience at 1.94 nMol/mL/min (reference range: 0.32 – 0.95 nMol/mL/min). Nagalase activity fell 27.8% after the three-month experience. Nagalase is a marker for inflammation often found elevated in viral infections, cancer and other chronic conditions (4,11,12). These results are consistent with previous observations (3,4) and can be interpreted as consequence of high levels of naturally formed Gc protein-derived Macrophage Activating Factor in the probiotic yogurt as hypothesized since 2011 (13) and elegantly demonstrated in 2020 (4).
Toxic metals in urine
Studies of toxic metals in urine, before and after intravenous (IV) chelation, demonstrated significant improvement associated with consumption of the probiotic yogurt.
Cadmium
Cadmium is a ubiquitous metal toxicant that can have adverse effects on the kidneys. The International Agency for Research on Cancer (IARC) of the World Health Organization (WHO) considers cadmium a Group 1 human carcinogen with prolonged exposure usually through the oral route (food or water). Cadmium is typically found in mining, paints, dyes, metallurgy, soils, and foods, but specifically mentioned is that it can enter rice through the soil and water. It is noted in the subject’s history that she consumes rice on a daily basis and has managed a garment factory for a number of years where she was exposed to dyes.
We previously demonstrated that cadmium may be responsible for the neurological signs and symptoms of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome and for abnormalities of breast cancer cell-induced angiogenesis (14, 15). In the subject, cadmium in the urine exceeded twice the maximum expected value at baseline in August. Pre-provocative (i.e., before IV chelation) baseline levels were high at 2.1 ug/g creatinine (reference interval high: <1 ug/g creatinine). Post-provocative cadmium remained high at 1.7 ug/g creatinine, using the IV chelating agent EDTA, as cadmium was being removed and is suggestive of a fair amount of the metal present in the body of the subject. In December, after completing the three-month experience with the probiotic yogurt, pre-provocative cadmium was 1.8 ug/g creatinine, still high and little changed from August; however, post-provocative cadmium was 2.4 ug/g creatinine, suggesting increased removal of cadmium. Such a removal could be explained
Blythe and Ruggiero 2020 – Probiotics and detoxification
4
considering that probiotics, such as those present in the product consumed by the subject, inhibit absorption of cadmium by protecting the intestinal barrier (16).
Lead
Lead levels pre-provocative in August were within a reference interval considered low risk at 0.5 ug/g creatinine. Post-provocative lead was high at 7.5 ug/g creatinine (reference range: < 2 ug/g creatinine), over 3 times the upper limit of the reference interval, suggesting the presence of substantial amounts of lead in the subject’s body that chelation was helping remove. In December, after the three-month experience with the probiotic yogurt, pre-provocative lead was again low risk at 0.7 ug/g creatinine, but post-provocation using a chelating agent, urine lead excretion significantly increased to 17 ug/g creatinine, which is 8.5 times the upper limit, considerably higher than in August and suggesting substantial elimination using both the probiotic yogurt and chelation. These results can be interpreted considering that probiotics, such as those present in the product consumed by the subject, are known to protect against lead toxicity (17) and may enhance lead elimination (18).
Barium
Barium is another toxic metal whose elimination was enhanced concomitant with consumption of the probiotic yogurt. This metal can interfere with calcium and potassium metabolism leading to hypokalemia with tingling of the extremities, loss of tendon reflexes and cardiac stimulation. Baseline pre-provocative barium in August was normal at 1.8 ug/g creatinine (reference range: <7 ug/g creatinine); post-provocative barium was 5.5 ug/g creatinine, suggesting some elimination of barium from the body with chelation. In December, after the three-month experience with the probiotic yogurt, pre-provocative barium remained normal at 3 ug/g creatinine. Post-provocative barium however was slightly elevated at 9.3 ug/g creatinine, suggesting further elimination. The subject remembered swallowing some material before an X-ray a number of years ago, presumably containing barium as a contrast agent, but could not fill in further details. Barium is also found in Brazil nuts, peanuts and peanut butter and certain fast foods, and is also found in seaweed and fish, all foods the subject eats. The working hypothesis is that, after the three-month nutritional program with the probiotic yogurt, barium was being eliminated. These results can be interpreted considering that removal of toxic metals is a common feature of probiotics, such as those present in the product consumed by the subject (19).
Non-metal toxicants in urine
Studies of non-metal toxicants in urine, before and after the three-month experience with the biodiverse probiotic yogurt, demonstrated significant changes for a number of toxicants as described in detail below.
Glyphosate
Blythe and Ruggiero 2020 – Probiotics and detoxification
5
Glyphosate is the world’s most widely produced herbicide and is found in runoff waters in agricultural areas. The WHO IARC considers glyphosate a probable Group 1 human carcinogen implicated in diseases as non-Hodgkin lymphoma, renal tubule carcinoma and other pathologies. Glyphosate disrupts the microbiome in the intestine, decreasing the ratio of beneficial to harmful microbes. Consistent with these effects of glyphosate, the subject’s stool sample showed a dysbiotic pattern with the absence of the usually present Lactobacilli strains.
Glyphosate in the subject’s urine showed a moderate presence at 1.35 ug/g creatinine in August. However, glyphosate fell to 0.8 ug/g creatinine, a 40% decline in three months, following the three-month experience with the probiotic yogurt. Beneficial Lactobacilli strains were believed added by the probiotic yogurt with the ratio of expected/beneficial flora registering a two-fold increase for Lactobacillus. These results can be interpreted considering that removal of non-metal toxicants is a well-established feature of probiotics, such as those present in the product consumed by the subject (3,20).
2-hydroxyisobutyric acid
2-hydroxyisobutyric acid (2HIB) in urine was within the reference values at 4,082 ug/g creatinine. 2HIB is an additive to improve vehicle octane ratings and is found in exhaust fumes. 2HIB has been shown to cause liver and kidney toxicity, and cancer in animals. After completing the three-month experience with the probiotic yogurt, urinary excretion of 2HIB rose 94% to 7,912 ug/g creatinine, thus suggesting elimination 2HIB from the body at an increased rate, consistent with well-established features of probiotics.
Perchlorate
Perchlorate (PERC) in urine in August was non-detectable; this result may be interpreted in two ways: a) the subject had not been exposed to PERC and no PERC had accumulated in her body; b) the subject was unable to excrete PERC. The first hypothesis seems less plausible because PERC is a ubiquitous pollutant. In addition to being used in rocket fuel and explosives, it is used in fertilizers and ordinary bleach, and often found contaminating food and water supplies. The second hypothesis was confirmed by the observation that, after completing the three-month experience with the probiotic yogurt, PERC was being excreted in December at 2.7 ug/g creatinine. Therefore, these results may be interpreted as increased excretion of PERC in association with consumption of the probiotic yogurt.
N-acetyl-S-(2-carbamoylethyl)cysteine
N-acetyl-S-(2-carbamoylethyl)cysteine (NAE) in urine was 22 ug/g creatinine in August. Acrylamide is the parent polymer used in industrial processes as plastics, food packaging, cosmetics and dyes. Foods like potato chips and French fries, and cigarette smoke exposure, are major sources. The toxicity occurs because asparagine, an amino acid important for central nervous system function, can produce acrylamide when cooked at high temperatures in the presence of sugars. High levels of acrylamide can elevate a patient’s risk of cancer and cause
Blythe and Ruggiero 2020 – Probiotics and detoxification
6
neurological damage. Foods rich in asparagine include asparagus, potatoes, legumes, nuts, seeds, beef, eggs and fish. After completing her three-month experience with the probiotic yogurt, NAE excretion levels increased substantially to 115 ug/g creatinine, well above the 75th percentile, indicating increased removal of this toxicant.
N-acetyl(propyl)cysteine
N-acetyl(propyl)cysteine (NAPR) in urine was moderately elevated in August at 19 ug/g creatinine, well above the 75th percentile. NAPR is an organic solvent and metabolite of 1- bromopropane used in dry cleaning, foam gluing and metal cleaning. It is a neurotoxin associated with sensory and motor deficits, decreased cognitive function, and with acute exposures, headaches. After completing the three-month experience with the probiotic yogurt in December, NAPR was no longer detectable, suggesting that substantial removal of the toxicant through urinary excretion had occurred.
2-hydroxyethyl mercapturic
A similar situation occurred with 2-hydroxyethyl mercapturic acid (HEMA), a toxicant that was detectable in the subject’s urine in August at 0.55 ug/g creatinine. HEMA comes from exposure to ethylene oxide used in agrochemicals, detergents, pharmaceuticals and personal care products. Ethylene oxide is also a sterilizer on rubber, plastics and electronics. Chronic exposure is mutagenic, and ethylene oxide is considered a carcinogen with increased incidence of breast cancer and leukemia. HEMA may also develop from exposure to vinyl chloride, an intermediate in the synthesis of these chemicals. These exposures have been associated with autism, headache, dizziness, liver damage and cancer. After completing the three-month experience with the probiotic yogurt in December, HEMA was no longer detectable, suggesting that substantial removal of the toxicant through urinary excretion had occurred.
Diethylphosphate
Diethylphosphate (DEP) in urine was moderately elevated in August at 3.7 ug/g creatinine. Organophosphates such as DEP are among the most toxic groups of substances in the world, and are primarily found in pesticide formulations. 85% of households in United States store at least one pesticide. These cholinesterase inhibitors cause sweating, salivation, diarrhea, and both depression and aggressive behavior. Autism spectrum disorder is associated with organophosphate exposure (21). It may be worth noting that the subject’s house was sprayed with pesticides in 2019. After completing the three-month experience with the probiotic yogurt in December, DEP was no longer detectable, suggesting that substantial removal of the toxicant through urinary excretion had occurred.
3-Phenoxybenzoic acid
3-Phenoxybenzoic acid (3BPA) in urine was moderately elevated in August at 2.1 ug/g creatinine, well above the 75th percentile. 3BPA is a metabolite of six different pyrethroid
Blythe and Ruggiero 2020 – Probiotics and detoxification
7
insecticides. They affect neurological development, disrupt hormones, induce cancers, and suppress the immune system. After completing the three-month experience with the probiotic yogurt in December, 3BPA was no longer detectable, suggesting that substantial removal of the toxicant through urinary excretion had occurred.
Stool analysis and microbiota status
Comprehensive stool analysis showed the absence of beneficial Lactobacillus strains at baseline in August. There were no parasites. Bacteria that are indicators of dysbiosis were present including Klebsiella oxytoca. As described above, toxicology testing found toxicants (Glyphosate) which disrupt the balance of the intestinal microbiota and may have contributed to the observed alterations. After completing the three-month experience with the probiotic yogurt in December, marked improvements were seen with the presence of Lactobacillus strains significantly repopulating the colon in high numbers with positive consequences for immune and other biological functions. Consistent with this observation, Klebsiella oxytoca, an indicator of dysbiosis, significantly decreased after completing the three-month experience.
As far as Short Chain Fatty Acids (SCFA) are concerned, significant results were observed for butyrate that increased 1.78 times (from 3.7% to 6.6% of total SCFA) after completing the three-month experience. It is worth noting that butyrate is produced by microbial fermentation in the colon of humans and animals. It is utilized as not only a primary nutrient that is the source of energy for epithelial cells of the colon, but also as a signaling molecule that regulates multiple functions of cells in the colon and other organs by optimizing gene expression, cell differentiation and tissue development, modulation of the immune system, and reduction of oxidative stress (22).
Discussion
Since this is an open-label, non-controlled, retrospective analysis, caution must be used when ascribing cause and effect to any nutritional approach outcome. However, the response associated with consumption of the probiotic yogurt was robust with regard to nagalase activity reduction, as well as to detoxification and microbiota reconstitution. Despite the short observational time period, the results were substantial and confirm and expand previous observations (3-6).
Reduction of serum nagalase activity is consistent with the observation that the extremely biodiverse probiotic yogurt described in this article has high Gc protein-derived Macrophage Activating Factor activity as demonstrated in recent studies (2,4). In addition to the effects on modulation of macrophage activity, it is worth noting that the probiotic yogurt used in this experience is highly fermented. It is known that the acidity associated with fermentation as well as the proteolytic activity of fermenting probiotics lead to formation of molecules, mainly derived from the hydrolysis of caseins, that have immune-modulatory and anti-oxidant properties (23). These may play an additional role in decreasing the subclinical inflammation associated with the presence of toxicants that in turn might have been responsible for the
Blythe and Ruggiero 2020 – Probiotics and detoxification
8
elevated nagalase activity observed prior to the experience described here. Also, the probiotic yogurt used in this study has the unique peculiarity of fermenting bovine colostrum; colostrum is endowed with intrinsic immune-modulating properties (24), and has high levels of vitamin D- binding protein that is the precursor of Gc protein-derived Macrophage Activating Factor (25).
Since nagalase is considered an indicator of cancer cell proliferation, viral infections as well as a marker of systemic inflammation (4,11, 12, 26), and since many of the toxicants described in this article are associated with increased cancer risk, immune dysfunction and chronic inflammation, it is tempting to speculate that decrease of nagalase activity represented a reduction of the risks associated with exposure to those toxicants. In other words, we hypothesize that there are two factors involved in the observed decrease of serum nagalase activity: on one side, the of Gc protein-derived Macrophage Activating Factor activity of the probiotic yogurt, a mechanism thoroughly described in previous studies (4,26); on the other side, the removal of toxicants that might have been responsible for elevated nagalase activity (3).
As far as the observed increased urinary excretion of toxic metals and other toxicants is concerned, the present observations extend and expand the results we first described in 2017 (3). In that article, we described increased urinary excretion of lipophilic toxicants associated with intermittent fasting, weight loss and consumption of the probiotic yogurt; in the present article, we are able to add that consumption of this peculiar probiotic yogurt is associated also with increased excretion of toxic metals and non-lipophilic toxicants such as Glyphosate. It is therefore plausible that the detoxifying effect of the probiotic yogurt is a general one and is not limited to a particular class of toxicants.
It is well assessed that probiotic strains help against metal toxicity by reducing the amount of toxicants in the liver and kidney, and by counteracting alterations in the levels of glutathione peroxidase and superoxide dismutase (27). In addition, probiotic strains may help eliminate toxicants by physical binding (28) as it is the case for lactic acid bacteria in the removal of Aflatoxin B1 (29). Furthermore, the present observations support the hypothesis that detoxification associated with probiotic yogurt consumption may be independent of mobilization of lipophilic toxicants from fat cells as a consequence of intermittent fasting or other weight loss protocols. In our previous article, we had postulated that mobilization of lipophilic toxicants from fat cells as a consequence of weight loss protocols was pre-requisite for their urinary excretion and, in that context, the probiotic yogurt helped by increasing the rate of urinary excretion (3). However, the subject described in this article did not lose any weight. Her body weight remained constant during the three-month experience. Therefore, we can deduce that consumption of the probiotic yogurt mentioned in this study helps with detoxification of all types of toxicants without the need of any further nutritional intervention.
The observed trend toward normalization of the gut microbiota and correction of dysbiosis is a well-known phenomenon associated with consumption of probiotics (30). In the case reported in this article, it may be argued that elimination of Glyphosate, a known disruptor of the healthy gut microbiota, contributed to the trend toward normalization.
Blythe and Ruggiero 2020 – Probiotics and detoxification
9
Conclusions
The experience of the subject suggests that the consumption of an extremely biodiverse probiotic yogurt described in references 1 and 2 is associated increased urinary excretion of toxicants, modulation of the immune system, and rebalance of the gut microbiota with overall positive effects on health. The authors wish to emphasize that the data in this case report are from one individual, and not part of a larger study even though they confirm and expand the results observed in other, unrelated, articles (3-6).
Without doubt, the observations described here can be classified as anecdotes. Anecdotes, or informal stories, once were the primary instrument for the advancement of medical knowledge; however, in recent times, anecdotes have less standing. According to this interpretation of the process of knowledge in medicine, controlled experiments such as randomized controlled trials provide medical knowledge while anecdotes do not (31). However, the authors share the convincing argument of the philosopher of science, Robert Nunn, when he challenged the deprecation of anecdotes in medicine since they contribute to the advancement of knowledge in medicine with the same significance of randomized controlled trials with impressive statistics or systematic reviews because “All of these stories become evidence of what works in medicine” (31).
Authors’ Contributions
Jerry Blythe, MD: observed the experience described in this article, provided critical input and assisted in revising and improving the article.
Marco Ruggiero, MD, PhD: wrote the first draft of this article, provided critical input and assisted in revising and improving the article.
Disclosures
Jerry Blythe discloses no conflict of interest. The subject described in this study bought all the foods and supplements used during her experience and paid for the analyses reported in this article.
Marco Ruggiero is the founder of Silver Spring Sagl, the company producing the probiotic yogurt used in this experience and has served as CEO of the company until his retirement in 2020. However, he had no prior knowledge of the nutritional plan followed by the subject of this article nor of the results of the analyses that were communicated by Dr. JB only after completion of the experience.
Ethics
Blythe and Ruggiero 2020 – Probiotics and detoxification
10
This article is original and contains material that has not been submitted or published in any other scientific journal.
References
1. Pacini S, Ruggiero M. Natural Plasmids in a Swiss Fermented Milk and Colostrum Product assessed by Microbiome Array. Madridge J Immunol. 2019; 3(2): 100-108. DOI: 10.18689/mjim- 1000123
2. Pacini S, Ruggiero M. Phage composition of a fermented milk and colostrum product assessed by microbiome array; putative role of open reading frames in reference to cell signaling and neurological development. J Neurol Stroke. 2020;10(2):80‒90. DOI: 10.15406/jnsk.2020.10.00416
3. Blythe J, Ruggiero M, Pacini S. Intermittent fasting and probiotic yogurt consumption are associated with reduction of serum alpha- N-acetylgalactosaminidase and increased urinary excretion of lipophilic toxicants. Madridge J Immunol. 2017; 1(1): 23-27. doi: 10.18689/mjim- 1000107
4. Carter M, Pacini S, Ruggiero M. Consumption of an Extremely Biodiverse Probiotic and a Supplement based on Microbial Chondroitin Sulfate is Associated with Very Low Serum Alpha- N- acetylgalactosaminidase (Nagalase) Activity and Decrease of C-reactive Protein Values. Am J Immunol 2020; 16: 8-18. DOI: 10.3844/ajisp.2020.8.18
5. Antonucci N, Pacini S, Ruggiero M. Use of an Extremely Biodiverse Probiotic and a Supplement Based on Microbial Chondroitin Sulfate is Associated with a Significant Decrease of Serum Free Kappa Light Chains as well as a Trend Toward Normalization of Kappa/Lambda Ratio and of Plasma Cell Bone Marrow Infiltration in a Case of Multiple Myeloma. Am J Immunol 2019; 15: 5-9. DOI: 10.3844/ajisp.2019.5.9
6. Zunaid IR, Pacini S, Ruggiero M. Significance of hydrophobic and charged sequence similarities in sodium-bile acid cotransporter and vitamin D-binding protein macrophage activating factor. bioRxiv 2020. DOI: https://doi.org/10.1101/2020.03.03.975524
7. Harben K, Schuchat A., Jenrnigan D et al. Transcript for CDC Update on Flu Activity. 2020. https://www.cdc.gov/media/releases/2018/t0202-flu-update-activity.html
8. John Hopkins Medicine. 102.3 Organization Policy on Single Case Reports and Case Series. “It is the policy of the Organization that a “single” case report (three or fewer cases) does not require review by the JHM IRB.“. https://www.hopkinsmedicine.org/institutional_review_board/guidelines_policies/organization _policies/102_3.html (accessed July 9, 2020)
Blythe and Ruggiero 2020 – Probiotics and detoxification
11
9. Cho YA, Kim J. Effect of Probiotics on Blood Lipid Concentrations: A Meta-Analysis of Randomized Controlled Trials. Medicine (Baltimore). 2015;94(43):e1714. doi:10.1097/MD.0000000000001714
10. Rizzoli, R., Biver, E. Are Probiotics the New Calcium and Vitamin D for Bone Health?. Curr Osteoporos Rep 18, 273–284 (2020). https://doi.org/10.1007/s11914-020-00591-6
11. Yamamoto, N, M. Urade, 2005. Pathogenic significance of alpha-N-acetylgalactosaminidase activity found in the hemagglutinin of influenza virus. Microbes Infect. Apr;7(4):674-81. DOI: 10.1016/j.micinf.2005.01.015
12. Yamamoto, N., 2006. Pathogenic significance of alpha-N-acetylgalactosaminidase activity found in the envelope glycoprotein gp160 of human immunodeficiency virus Type 1. AIDS Res Hum Retroviruses. Mar;22(3):262-71. DOI: 10.1089/aid.2006.22.262
13. Pacini S, Punzi T, Morucci G, Ruggiero M. Macrophages of the mucosa- associated lymphoid tissue (MALT) as key elements of the immune response to vitamin d binding protein- macrophage activating factor. It J Anat Embryol. 2011; 116(1): 136. doi: 10.13128/IJAE-10160 https://www.torrossa.com/en/catalog/preview/2490419
14. Pacini S, Fiore MG, Magherini S, et al. Could cadmium be responsible for some of the neurological signs and symptoms of Myalgic Encephalomyelitis/Chronic Fatigue Syndrome. Med Hypotheses. 2012;79(3):403-407. doi:10.1016/j.mehy.2012.06.007
15. Pacini S, Punzi T, Morucci G, Gulisano M, Ruggiero M. A paradox of cadmium: a carcinogen that impairs the capability of human breast cancer cells to induce angiogenesis. J Environ Pathol Toxicol Oncol. 2009;28(1):85-88.
16. Zhai Q, Tian F, Zhao J, Zhang H, Narbad A, Chen W. Oral Administration of Probiotics Inhibits Absorption of the Heavy Metal Cadmium by Protecting the Intestinal Barrier. Appl Environ Microbiol. 2016;82(14):4429-4440. Published 2016 Jun 30. doi:10.1128/AEM.00695-16
17. Zhai Q, Yang L, Zhao J, Zhang H, Tian F, Chen W. Protective Effects of Dietary Supplements Containing Probiotics, Micronutrients, and Plant Extracts Against Lead Toxicity in Mice. Front Microbiol. 2018;9:2134. Published 2018 Sep 11. doi:10.3389/fmicb.2018.02134
18.
J Microbiol
Yi YJ, Lim JM, Gu S, et al. Potential use of lactic acid bacteria Leuconostoc mesenteroides as

a probiotic for the removal of Pb(II) toxicity.
19. Monachese M, Burton JP, Reid G. Bioremediation and tolerance of humans to heavy metals through microbial processes: a potential role for probiotics? Appl Environ Microbiol. 2012;78(18):6397-6404. doi:10.1128/AEM.01665-12
Blythe and Ruggiero 2020 – Probiotics and detoxification
12
. 2017;55(4):296-303.
doi:10.1007/s12275-017-6642-x
20. He, F., Zuo, L., Ward, E., and Arciero, P.J. Serum polychlorinated biphenyls increase and oxidative stress decreases with a protein-pacing caloric restriction diet in obese men and women. Int. J. Environ. Res. Public Health; 2017 14: 59. doi:10.3390/ijerph14010059
21. Sagiv SK, Harris MH, Gunier RB, et al. Prenatal Organophosphate Pesticide Exposure and Traits Related to Autism Spectrum Disorders in a Population Living in Proximity to Agriculture [published correction appears in Environ Health Perspect. 2018 Jul 05;126(7):079001]. Environ Health Perspect. 2018;126(4):047012. Published 2018 Apr 25. doi:10.1289/EHP2580
22. Bedford A, Gong J. Implications of butyrate and its derivatives for gut health and animal production. Anim Nutr. 2018;4(2):151-159. doi:10.1016/j.aninu.2017.08.010
23. Ebner, J., Aşçı Arslan, A., Fedorova, M., Hoffmann, R., Küçükçetin, A. and Pischetsrieder, M. Peptide profiling of bovine kefir reveals 236 unique peptides released from caseins during its production by starter culture or kefir grains. J. Proteomics. 2015; 117:41-57. doi: 10.1016/j.jprot.2015.01.005
24. Shing, C.M., Hunter, D.C. and Stevenson, L.M. Bovine colostrum supplementation and exercise performance: potential mechanisms. Sports Med. 2009; 39:1033-1054. doi:10.2165/11317860-000000000-00000
25. Senda, A., Fukuda, K., Ishii, T. and Urashima, T. Changes in the bovine whey proteome during the early lactation period. Anim. Sci J. 2011; 82:698-706. doi:10.1111/j.1740-0929.2011.00886.x
26. Thyer, L., Ward, E., Smith, R., et al. GC protein-derived macrophage-activating factor decreases α-N-acetylgalactosaminidase levels in advanced cancer patients. Oncoimmunology. 2013. 2:e25769. doi: 10.4161/onci.25769
27. Majlesi, M., Shekarforoush, S.S., Ghaisari, H.R., Nazifi, S., Sajedianfard, J. and Eskandari, M.H. Effect of probiotic Bacillus coagulans and Lactobacillus plantarum on alleviation of mercury toxicity in rat. Probiotics Antimicrob. Proteins. 2017; Jan 13. doi:10.1007/s12602-016- 9250-x
28. Zoghi, A., Khosravi-Darani, K. and Sohrabvandi, S. Surface binding of toxins and heavy metals by probiotics. Mini Reviews in Med. Chem. 2014; 14:84-98. doi:10.2174/1389557513666131211105554
29. Haskard, C.A., El Nezami, H. S., Kankaanpää, P. E., Salminen, S. and Ahokas, J.T.
Surface binding of Aflatoxin B1 by lactic acid bacteria. Appl. Environ. Microbiol. 2001; 67:3086– 3091. doi: 10.1128/AEM.67.7.3086–3091.2001
Blythe and Ruggiero 2020 – Probiotics and detoxification
13
30. Bull MJ, Plummer NT. Part 2: Treatments for Chronic Gastrointestinal Disease and Gut Dysbiosis. Integr Med (Encinitas). 2015 Feb;14(1):25-33. PMID: 26770128; PMCID: PMC4566455.
31. Nunn R. Mere anecdote: evidence and stories in medicine. J Eval Clin Pract. 2011;17(5):920- 926. doi:10.1111/j.1365-2753.2011.01727.x



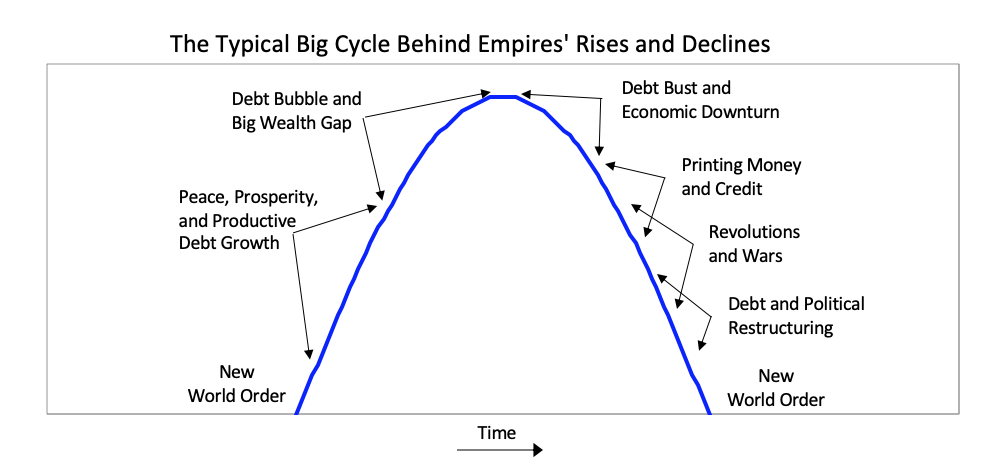 In brief, after the creation of a new set of rules establishes the new world order, there is typically a peaceful and prosperous period. As people get used to this they increasingly bet on the prosperity continuing, and they increasingly borrow money to do that, which eventually leads to a bubble. As the prosperity increases the wealth gap grows. Eventually the debt bubble bursts, which leads to the printing of money and credit and increased internal conflict, which leads to some sort of wealth redistribution revolution that can be peaceful or violent. Typically at that time late in the cycle the leading empire that won the last economic and geopolitical war is less powerful relative to rival powers that prospered during the prosperous period, and with the bad economic conditions and the disagreements between powers there is typically some kind of war. Out of these debt, economic, domestic, and world-order breakdowns that take the forms of revolutions and wars come new winners and losers. Then the winners get together to create the new domestic and world orders. That is what has repeatedly happened through time. The lines in the chart signify the relative powers of the 11 most powerful empires over the last 500 years. In the chart below you can see where the US and China are currently in their cycles. As you can see the United States is now the most powerful empire by not much, it is in relative decline, Chinese power is rapidly rising, and no other powers come close.
In brief, after the creation of a new set of rules establishes the new world order, there is typically a peaceful and prosperous period. As people get used to this they increasingly bet on the prosperity continuing, and they increasingly borrow money to do that, which eventually leads to a bubble. As the prosperity increases the wealth gap grows. Eventually the debt bubble bursts, which leads to the printing of money and credit and increased internal conflict, which leads to some sort of wealth redistribution revolution that can be peaceful or violent. Typically at that time late in the cycle the leading empire that won the last economic and geopolitical war is less powerful relative to rival powers that prospered during the prosperous period, and with the bad economic conditions and the disagreements between powers there is typically some kind of war. Out of these debt, economic, domestic, and world-order breakdowns that take the forms of revolutions and wars come new winners and losers. Then the winners get together to create the new domestic and world orders. That is what has repeatedly happened through time. The lines in the chart signify the relative powers of the 11 most powerful empires over the last 500 years. In the chart below you can see where the US and China are currently in their cycles. As you can see the United States is now the most powerful empire by not much, it is in relative decline, Chinese power is rapidly rising, and no other powers come close. 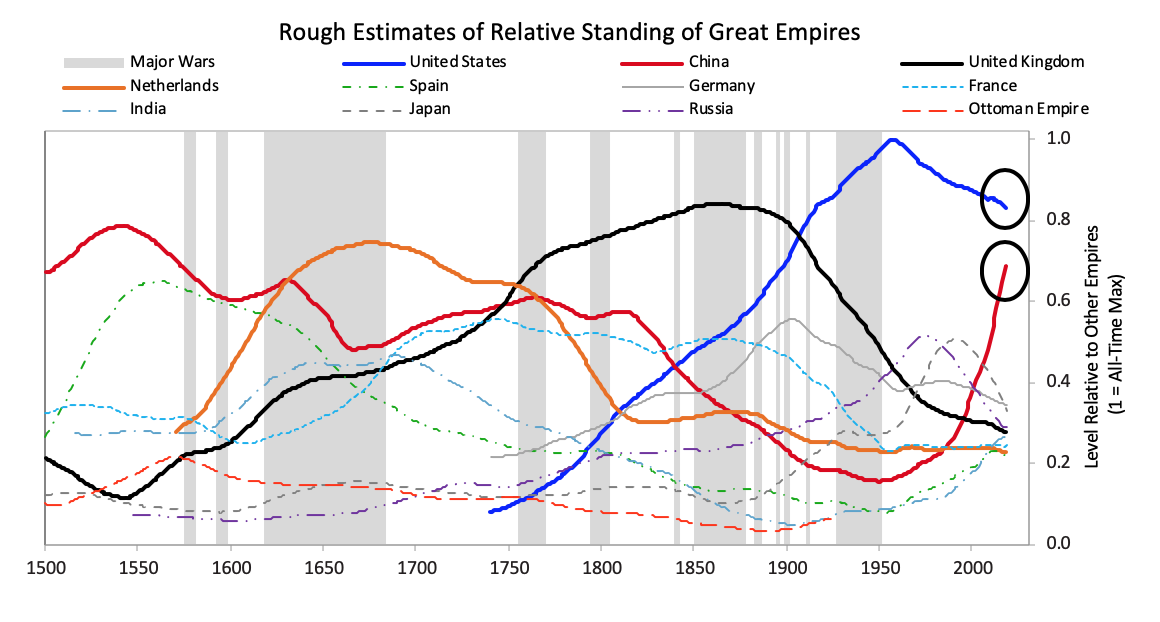 Because that chart is a bit confusing, for simplicity the next chart shows the same lines as in that chart except for just the most powerful reserve currency empires (which are based on an average of eight different measures of power that we explained in Chapter 1 and will explore more carefully in this chapter).
Because that chart is a bit confusing, for simplicity the next chart shows the same lines as in that chart except for just the most powerful reserve currency empires (which are based on an average of eight different measures of power that we explained in Chapter 1 and will explore more carefully in this chapter).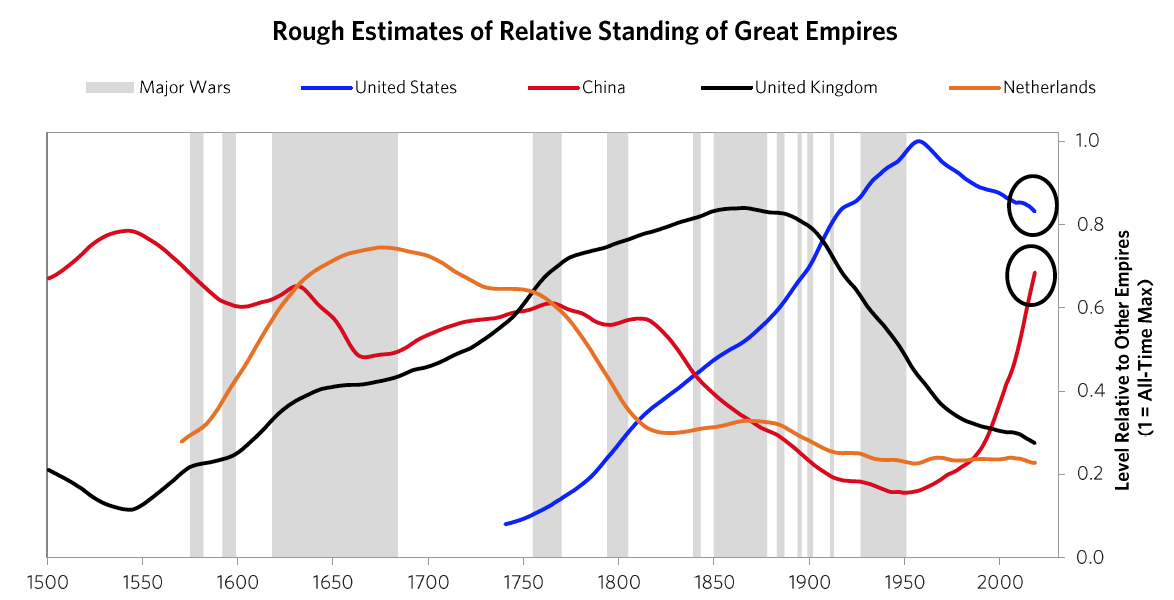 The next chart offers an even more simplified view. As shown, the United States and China are the only two major powers, you can see where each of their Big Cycles is, and you can see that they are approaching comparability, which is when the risks of wars of one type or another are greater than when the leading powers are earlier in the cycle. To be clear, I didn’t start out trying to make an argument and then go looking for stats to support it; doing that doesn’t work in my profession as only accuracy pays. I simply gathered stats that reflected these different measures of strength and put them in these indices, which led to these results. I suspect that if you did that exercise yourself picking whatever stats you’d like you’d see a similar picture, and I suspect that what I’m showing you here rings true to you if you’re paying attention to such things. For those reasons I suspect that all I am doing is helping you put where we are in perspective. To reiterate, I am not saying anything about the future. I will do that in the concluding chapter of this book. All I want to do is bring you up to date and, in the process, make clear how these cycles have worked in the past, which will also alert you to the markers to watch out for and help you see where in the cycles the major countries are and what is likely to come next.
The next chart offers an even more simplified view. As shown, the United States and China are the only two major powers, you can see where each of their Big Cycles is, and you can see that they are approaching comparability, which is when the risks of wars of one type or another are greater than when the leading powers are earlier in the cycle. To be clear, I didn’t start out trying to make an argument and then go looking for stats to support it; doing that doesn’t work in my profession as only accuracy pays. I simply gathered stats that reflected these different measures of strength and put them in these indices, which led to these results. I suspect that if you did that exercise yourself picking whatever stats you’d like you’d see a similar picture, and I suspect that what I’m showing you here rings true to you if you’re paying attention to such things. For those reasons I suspect that all I am doing is helping you put where we are in perspective. To reiterate, I am not saying anything about the future. I will do that in the concluding chapter of this book. All I want to do is bring you up to date and, in the process, make clear how these cycles have worked in the past, which will also alert you to the markers to watch out for and help you see where in the cycles the major countries are and what is likely to come next.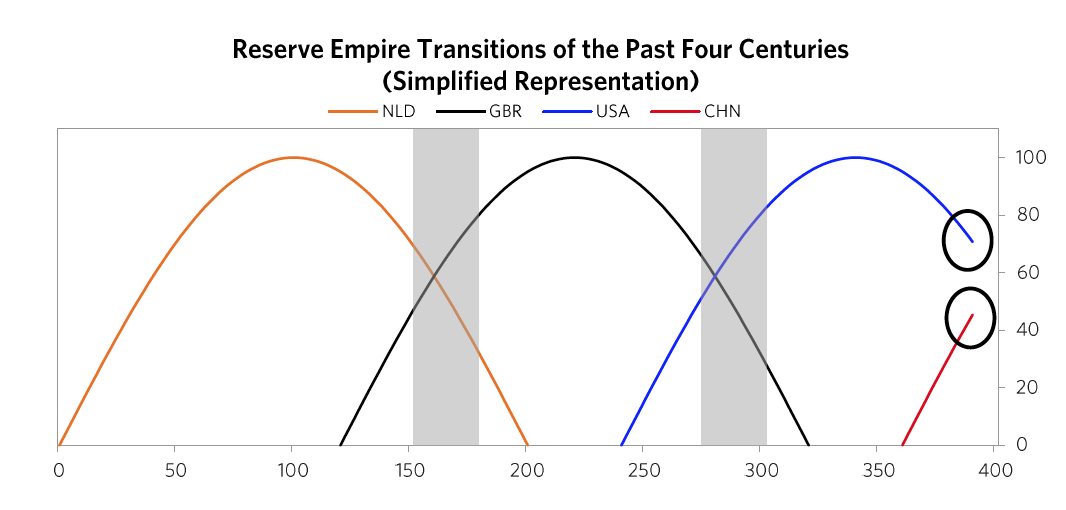 The chart below from Chapter 1 shows this play out via the eight measures of strength—education, innovation and technology, competitiveness, military, trade, output, financial center, and reserve status—that we capture in the aggregate charts. It shows the average of each of these measures of strength, with most of the weight on the most recent three reserve countries (the US, the UK, and the Dutch).
The chart below from Chapter 1 shows this play out via the eight measures of strength—education, innovation and technology, competitiveness, military, trade, output, financial center, and reserve status—that we capture in the aggregate charts. It shows the average of each of these measures of strength, with most of the weight on the most recent three reserve countries (the US, the UK, and the Dutch).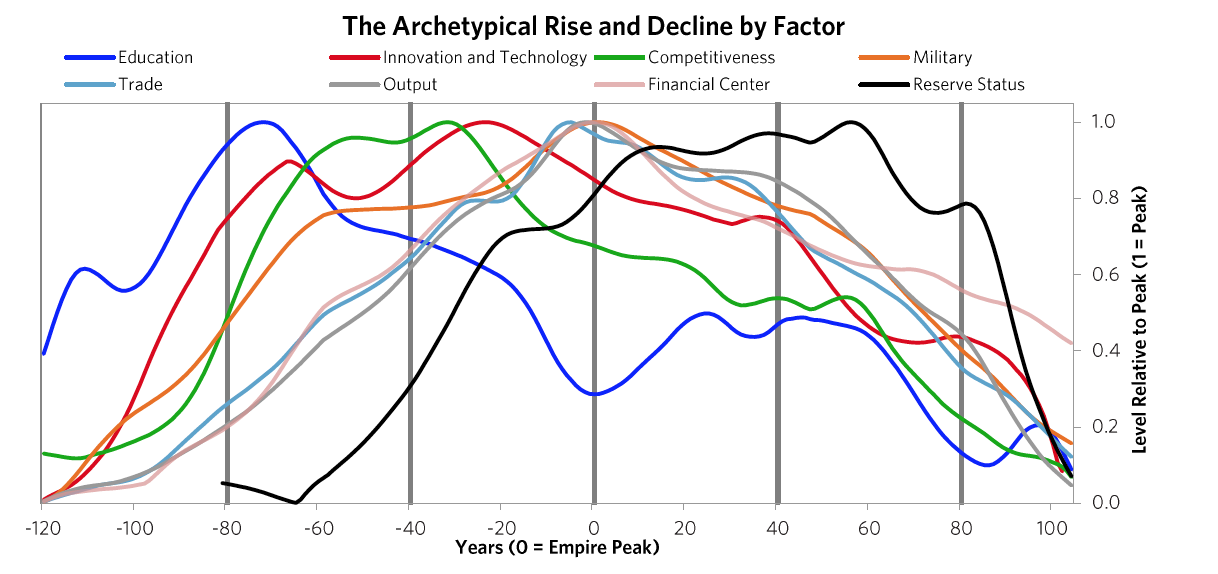 As explained in Chapter 1, in brief these strengths and weaknesses are mutually reinforcing—i.e., strengths and weaknesses in education, competitiveness, economic output, share of world trade, etc., contribute to the others being strong or weak, for logical reasons—and their order is broadly indicative of the processes that lead to the rising and declining of empires. For example, quality of education has been the long-leading strength of rises and declines in these measures of power, and the long-lagging strength has been the reserve currency. That is because strong education leads to strengths in most areas, including the creation of the world’s most common currency. That common currency, just like the world’s common language, tends to stay around because the habit of usage lasts longer than the strengths that made it so commonly used. We will now look at the specifics more closely, starting with how these Big Cycles have played out over the last 500 years and then looking at the declines of the Dutch and British empires so you can see how these things go.1) The Last 500 Years in About 4,000 Words The Rise & Decline of the Dutch Empire and the Dutch Guilder In the 1500-1600 period the Spanish empire was the pre-eminent economic empire in the “Western” world while the Chinese empire under the Ming Dynasty was the most powerful empire in the “Eastern” world, even more powerful than the Spanish empire (see the green dashed line and the red solid line in chart 2). The Spanish got rich by taking their ships and military power around the world, seizing control of vast areas (13% of the landmass of the earth!) and extracting valuable things from them, most importantly gold and silver which were the money of the time. As shown by the orange line in the chart of the relative standing of the great empires, the Dutch gained power as Spanish power was waning. At the time Spain controlled the small area we now call Holland. When the Dutch became powerful enough in 1581, they overthrew the Spanish and went on to eclipse both the Spanish and the Chinese as the world’s richest empire from around 1625 to their collapse in 1780. The Dutch empire reached its peak around 1650 in what was called the Dutch Golden Age. This period was one of great globalization as ships that could travel around the world to gain the riches that were out there flourished, and the Dutch, with their great shipbuilding and their economic system, were ahead of others in using ships, economic rewards, and military power to build their empire. Holland (as we now call it) remained the richest power for about 100 years. How did that happen?The Dutch were superbly educated people who were very inventive—in fact they came up with 25% of all major inventions in the world at their peak in the 17th century. The two most important inventions they came up with were 1) ships that were uniquely good that could take them all around the world, which, with the military skills that they acquired from all the fighting they did in Europe, allowed them to collect great riches around the world, and 2) the capitalism that fueled these endeavors. Not only did the Dutch follow a capitalist approach to resource allocation, they invented capitalism. By capitalism I mean public debt and equity markets. Of course production existed before, but that is not capitalism, and of course trade existed before, but that is not capitalism, and of course private ownership existed before, but that is not capitalism. By capitalism I mean the ability of large numbers of people to collectively lend money and buy ownership in money-making endeavors. The Dutch created that when they invented the first listed public company (the Dutch East India Company) and the first stock exchange in 1602 and when they built the first well-developed lending system in which debt could more easily be created. They also created the world’s first reserve currency. The Dutch guilder was the first “world reserve currency” other than gold and silver because it was the first empire to extend around much of the world and to have its currency so broadly accepted. Fueled by these qualities and strengths, the Dutch empire continued to rise on a relative basis until around 1700 when the British started to grow strongly. The numerous investment market innovations of the Dutch and their successes in producing profits attracted investors, which led to Amsterdam becoming the world’s leading financial center; the Dutch government channeled money into debt and some equity investments in various businesses, the most important of which was the Dutch East India Company.At this time of prosperity, other countries grew in power too. As other countries became more competitive, the Dutch empire became more costly and less competitive, and it found maintaining its empire less profitable and more challenging. Most importantly the British got stronger economically and militarily in the classic ways laid out in Chapter 1. Before they had become clear competitors they had military partnerships during most of the 80+ years leading up to the Fourth Anglo-Dutch War. That changed over time as they bumped into each other in the same markets. The Dutch and British had lots of conflicts over economic issues. For example, the English made a law that only English ships could be used to import goods into England, which hurt Dutch shipping companies that had a big business of shipping others’ goods to England, which led to the English seizing Dutch ships and expanding the British East India Company. Typically before all-out war is declared there is about a decade of these sorts of economic, technological, geopolitical, and capital wars when the conflicting powers approach comparability and test and try to intimidate each other’s powers. At the time the British came up with military inventions and built more naval strength, and they continued to gain relative economic strength. As shown in the chart of relative standing of empires shown above, around 1750 the British became a stronger power than the Dutch, particularly economically and militarily, both because the British (and French) became stronger and because the Dutch became weaker. As is classic the Dutch a) became more indebted, b) had a lot of internal fighting over wealth (between its states/provinces, between the rich and the poor, and between political factions)
As explained in Chapter 1, in brief these strengths and weaknesses are mutually reinforcing—i.e., strengths and weaknesses in education, competitiveness, economic output, share of world trade, etc., contribute to the others being strong or weak, for logical reasons—and their order is broadly indicative of the processes that lead to the rising and declining of empires. For example, quality of education has been the long-leading strength of rises and declines in these measures of power, and the long-lagging strength has been the reserve currency. That is because strong education leads to strengths in most areas, including the creation of the world’s most common currency. That common currency, just like the world’s common language, tends to stay around because the habit of usage lasts longer than the strengths that made it so commonly used. We will now look at the specifics more closely, starting with how these Big Cycles have played out over the last 500 years and then looking at the declines of the Dutch and British empires so you can see how these things go.1) The Last 500 Years in About 4,000 Words The Rise & Decline of the Dutch Empire and the Dutch Guilder In the 1500-1600 period the Spanish empire was the pre-eminent economic empire in the “Western” world while the Chinese empire under the Ming Dynasty was the most powerful empire in the “Eastern” world, even more powerful than the Spanish empire (see the green dashed line and the red solid line in chart 2). The Spanish got rich by taking their ships and military power around the world, seizing control of vast areas (13% of the landmass of the earth!) and extracting valuable things from them, most importantly gold and silver which were the money of the time. As shown by the orange line in the chart of the relative standing of the great empires, the Dutch gained power as Spanish power was waning. At the time Spain controlled the small area we now call Holland. When the Dutch became powerful enough in 1581, they overthrew the Spanish and went on to eclipse both the Spanish and the Chinese as the world’s richest empire from around 1625 to their collapse in 1780. The Dutch empire reached its peak around 1650 in what was called the Dutch Golden Age. This period was one of great globalization as ships that could travel around the world to gain the riches that were out there flourished, and the Dutch, with their great shipbuilding and their economic system, were ahead of others in using ships, economic rewards, and military power to build their empire. Holland (as we now call it) remained the richest power for about 100 years. How did that happen?The Dutch were superbly educated people who were very inventive—in fact they came up with 25% of all major inventions in the world at their peak in the 17th century. The two most important inventions they came up with were 1) ships that were uniquely good that could take them all around the world, which, with the military skills that they acquired from all the fighting they did in Europe, allowed them to collect great riches around the world, and 2) the capitalism that fueled these endeavors. Not only did the Dutch follow a capitalist approach to resource allocation, they invented capitalism. By capitalism I mean public debt and equity markets. Of course production existed before, but that is not capitalism, and of course trade existed before, but that is not capitalism, and of course private ownership existed before, but that is not capitalism. By capitalism I mean the ability of large numbers of people to collectively lend money and buy ownership in money-making endeavors. The Dutch created that when they invented the first listed public company (the Dutch East India Company) and the first stock exchange in 1602 and when they built the first well-developed lending system in which debt could more easily be created. They also created the world’s first reserve currency. The Dutch guilder was the first “world reserve currency” other than gold and silver because it was the first empire to extend around much of the world and to have its currency so broadly accepted. Fueled by these qualities and strengths, the Dutch empire continued to rise on a relative basis until around 1700 when the British started to grow strongly. The numerous investment market innovations of the Dutch and their successes in producing profits attracted investors, which led to Amsterdam becoming the world’s leading financial center; the Dutch government channeled money into debt and some equity investments in various businesses, the most important of which was the Dutch East India Company.At this time of prosperity, other countries grew in power too. As other countries became more competitive, the Dutch empire became more costly and less competitive, and it found maintaining its empire less profitable and more challenging. Most importantly the British got stronger economically and militarily in the classic ways laid out in Chapter 1. Before they had become clear competitors they had military partnerships during most of the 80+ years leading up to the Fourth Anglo-Dutch War. That changed over time as they bumped into each other in the same markets. The Dutch and British had lots of conflicts over economic issues. For example, the English made a law that only English ships could be used to import goods into England, which hurt Dutch shipping companies that had a big business of shipping others’ goods to England, which led to the English seizing Dutch ships and expanding the British East India Company. Typically before all-out war is declared there is about a decade of these sorts of economic, technological, geopolitical, and capital wars when the conflicting powers approach comparability and test and try to intimidate each other’s powers. At the time the British came up with military inventions and built more naval strength, and they continued to gain relative economic strength. As shown in the chart of relative standing of empires shown above, around 1750 the British became a stronger power than the Dutch, particularly economically and militarily, both because the British (and French) became stronger and because the Dutch became weaker. As is classic the Dutch a) became more indebted, b) had a lot of internal fighting over wealth (between its states/provinces, between the rich and the poor, and between political factions)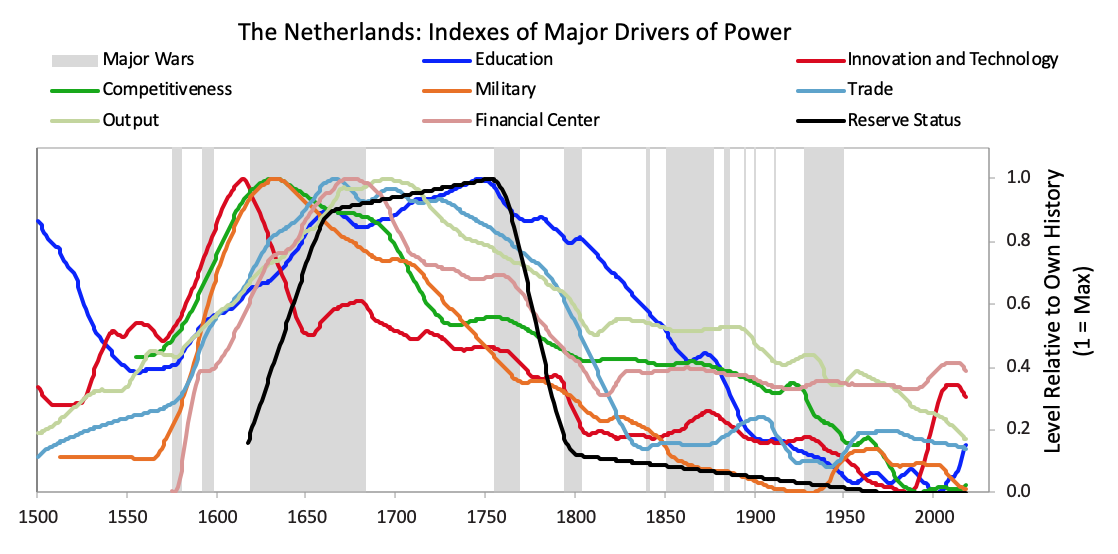 After declaring independence in 1581, the Dutch fought off the Spanish and built a global trading empire that became responsible for over a third of global trade largely via the first mega-corporation, the Dutch East India Company. As shown in the chart above, with a strong educational background the Dutch innovated in a number of areas. They produced roughly 25% of global inventions in the early 17th century,
After declaring independence in 1581, the Dutch fought off the Spanish and built a global trading empire that became responsible for over a third of global trade largely via the first mega-corporation, the Dutch East India Company. As shown in the chart above, with a strong educational background the Dutch innovated in a number of areas. They produced roughly 25% of global inventions in the early 17th century,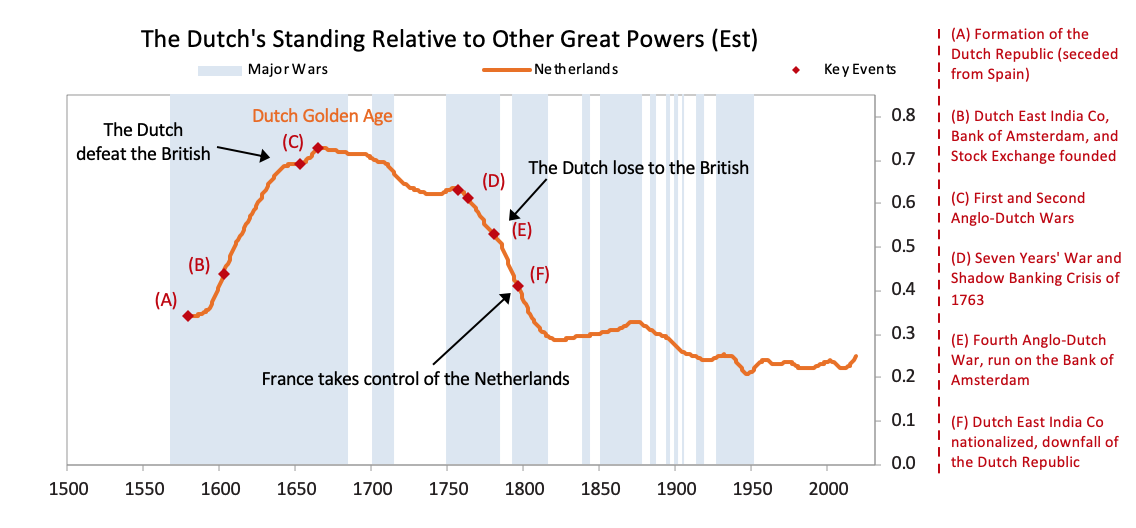 As shown, the seeds of Dutch decline were sown in the latter part of the 17th century as they started to lose their competitiveness and became overextended globally trying to support an empire that had become more costly than profitable. Increased debt-service payments squeezed them while their worsening competitiveness hurt their income from trade. Earnings from business abroad also fell. Wealthy Dutch savers moved their cash abroad both to get out of Dutch investments and into British investments, which were more attractive due to strong earnings growth and higher yields.
As shown, the seeds of Dutch decline were sown in the latter part of the 17th century as they started to lose their competitiveness and became overextended globally trying to support an empire that had become more costly than profitable. Increased debt-service payments squeezed them while their worsening competitiveness hurt their income from trade. Earnings from business abroad also fell. Wealthy Dutch savers moved their cash abroad both to get out of Dutch investments and into British investments, which were more attractive due to strong earnings growth and higher yields.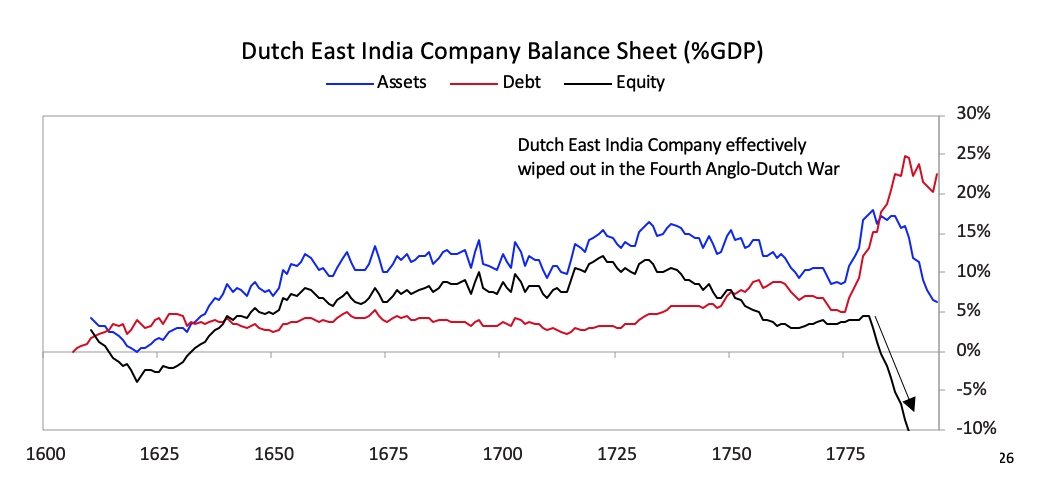 [26]As shown in the chart below the Dutch East India Company, which was essentially the Dutch economy and military wrapped into a company, started to make losses in 1780, which became enormous during the Fourth Anglo-Dutch War.
[26]As shown in the chart below the Dutch East India Company, which was essentially the Dutch economy and military wrapped into a company, started to make losses in 1780, which became enormous during the Fourth Anglo-Dutch War. 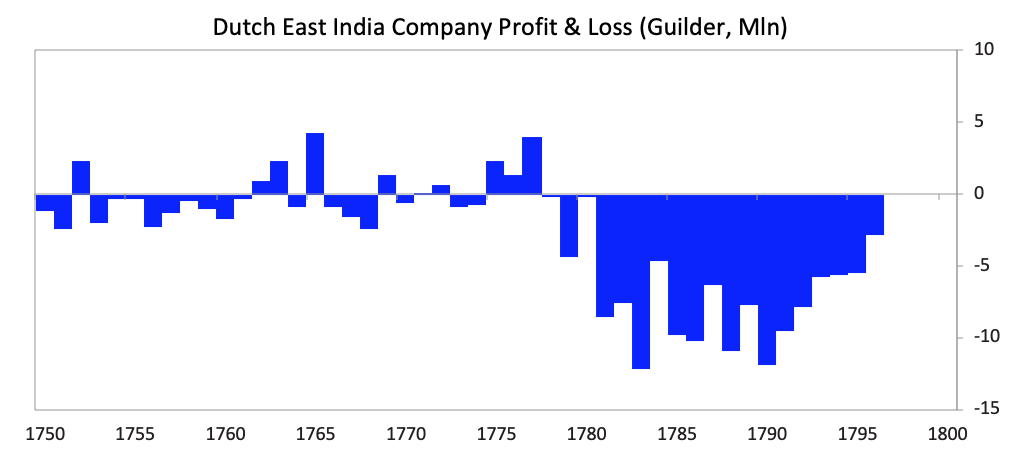 As deposit holders at the Bank of Amsterdam realized the bank was “lending” freshly printed guilders to save the Dutch East India Company, there was a run on the Bank of Amsterdam.
As deposit holders at the Bank of Amsterdam realized the bank was “lending” freshly printed guilders to save the Dutch East India Company, there was a run on the Bank of Amsterdam.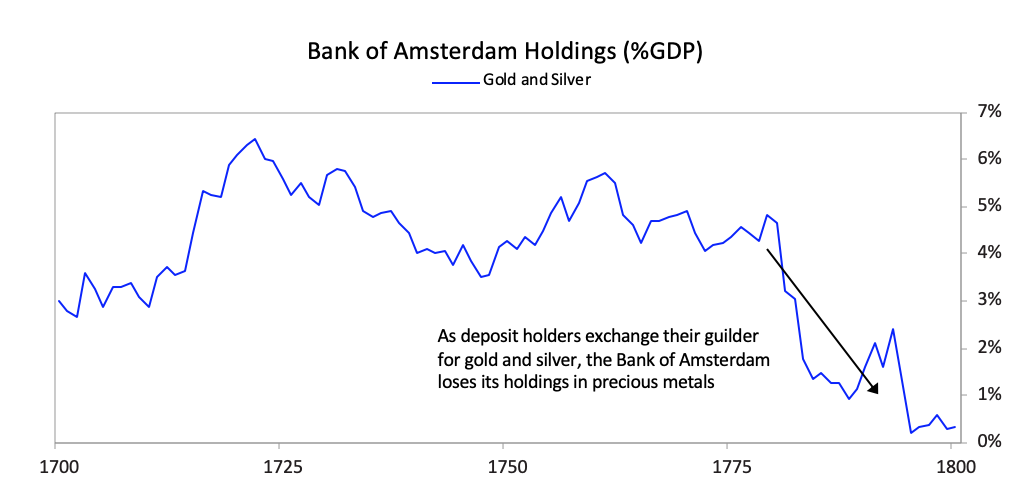 The Bank of Amsterdam had no choice since the company was too important to allow to fail both because of its significance to the economy and its outstanding debt in the Dutch financial system, so the Bank of Amsterdam began “lending” large sums of newly printed guilders to the company. During the war, policy makers also used the bank to lend to the government.
The Bank of Amsterdam had no choice since the company was too important to allow to fail both because of its significance to the economy and its outstanding debt in the Dutch financial system, so the Bank of Amsterdam began “lending” large sums of newly printed guilders to the company. During the war, policy makers also used the bank to lend to the government.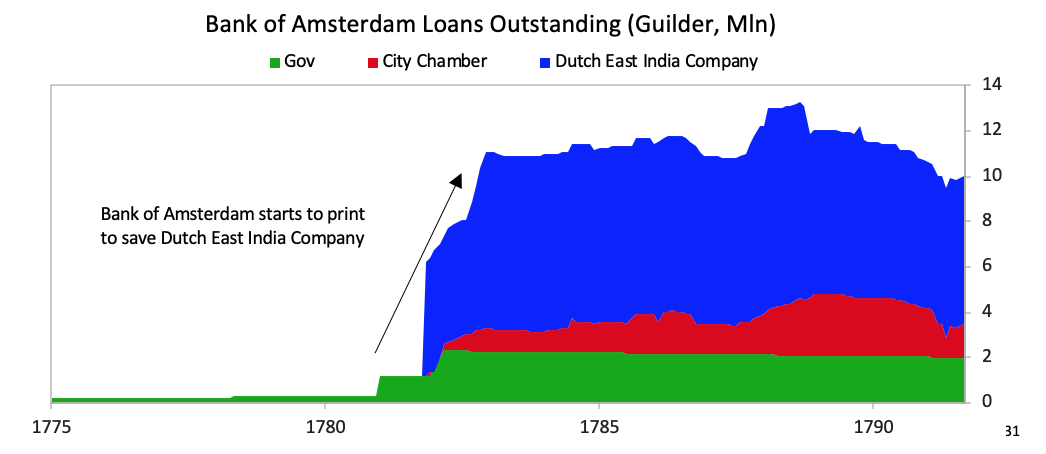 [31]Interest rates rose and the Bank of Amsterdam had to devalue, undermining the credibility of the guilder as a storehold of value.
[31]Interest rates rose and the Bank of Amsterdam had to devalue, undermining the credibility of the guilder as a storehold of value.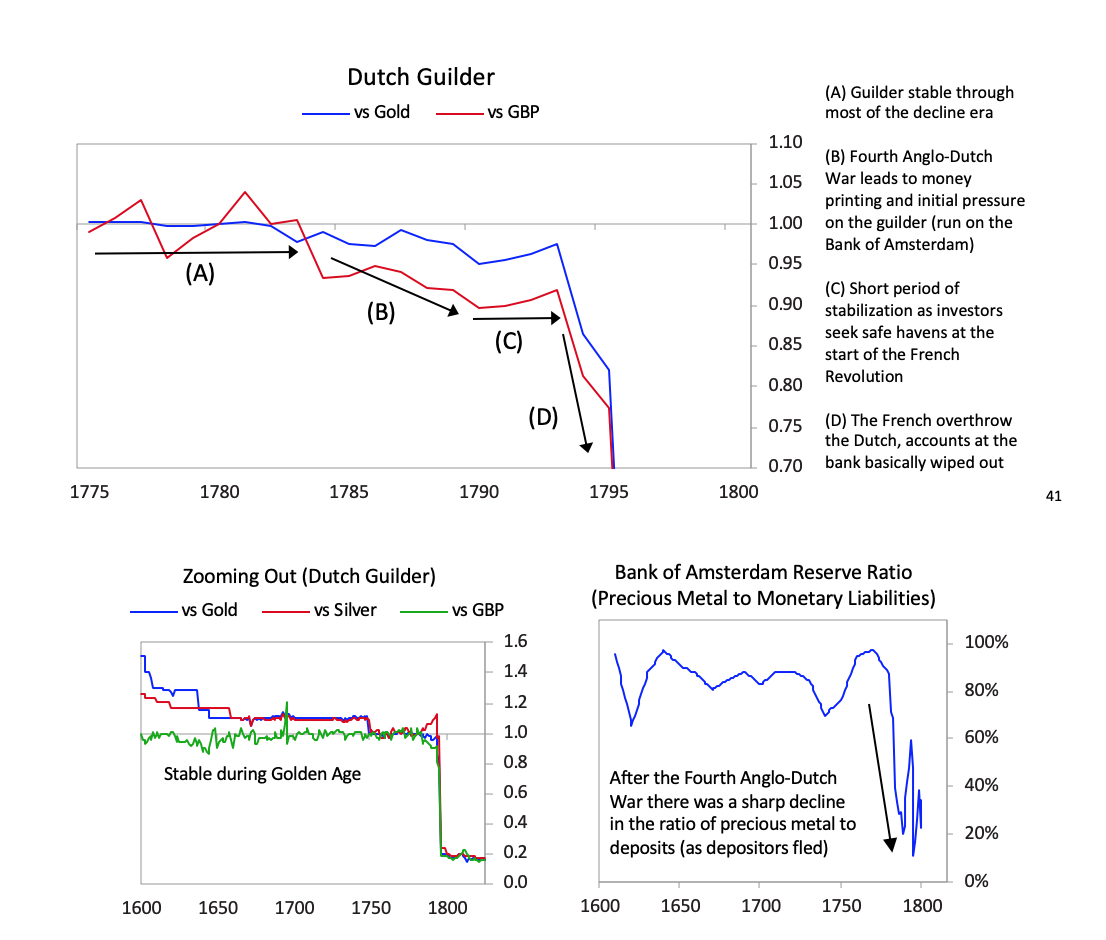 [41]The chart below shows the returns of holding the Dutch East India Company for investors starting in various years. As with most bubble companies, it originally did great, with great fundamentals, which attracted more investors even as its fundamentals started to weaken, but it increasingly got into debt, until the failed fundamentals and excessive debt burdens broke the company.
[41]The chart below shows the returns of holding the Dutch East India Company for investors starting in various years. As with most bubble companies, it originally did great, with great fundamentals, which attracted more investors even as its fundamentals started to weaken, but it increasingly got into debt, until the failed fundamentals and excessive debt burdens broke the company. 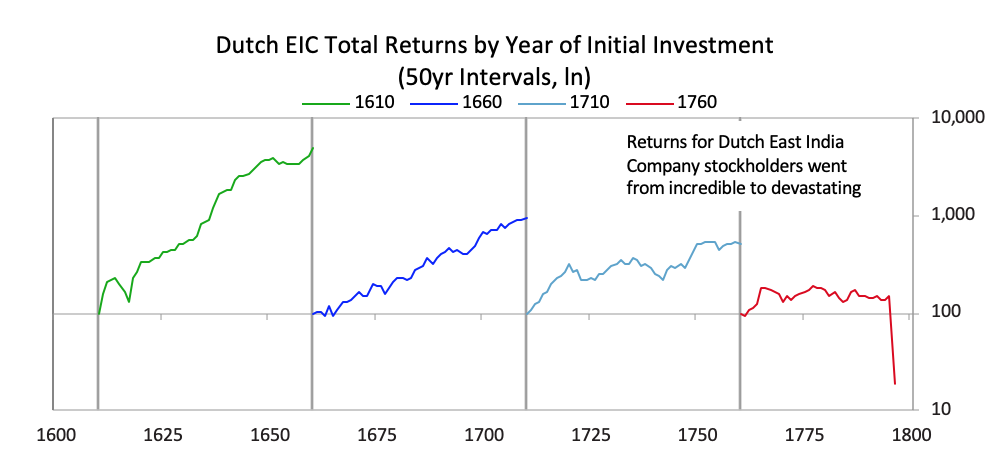 As is typical, with the decline in power of the leading empire and the rise in power of the new empire, the returns of investment assets in the declining empire fell relative to the returns of investing in the rising empire. For example, as shown below, the returns on investments in the British East India Company far exceeded those in the Dutch East India Company, and the returns of investing in Dutch government bonds were terrible relative to the returns of investing in English government bonds. This was reflective of virtually all investments in these two countries.
As is typical, with the decline in power of the leading empire and the rise in power of the new empire, the returns of investment assets in the declining empire fell relative to the returns of investing in the rising empire. For example, as shown below, the returns on investments in the British East India Company far exceeded those in the Dutch East India Company, and the returns of investing in Dutch government bonds were terrible relative to the returns of investing in English government bonds. This was reflective of virtually all investments in these two countries. 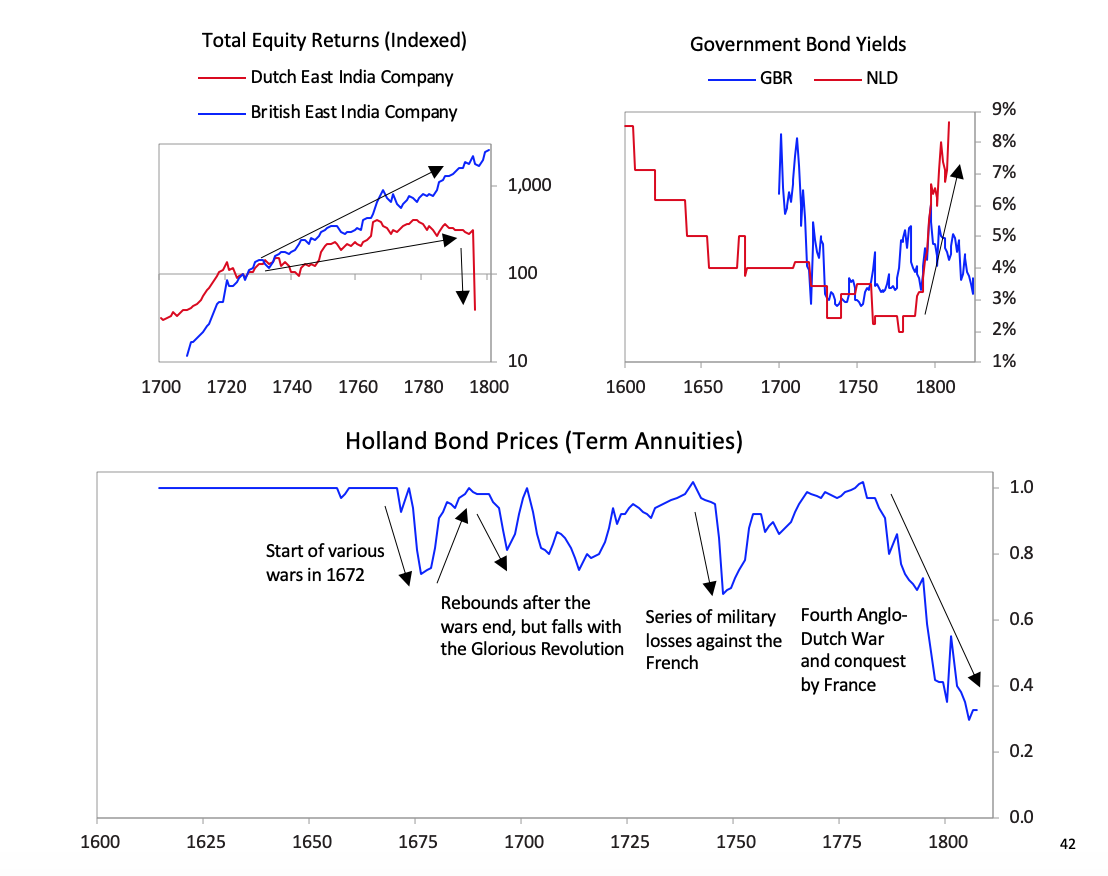 [42]The British Empire and the British PoundBefore we get to the collapse of the British empire and the British pound, let’s take a quick look at the whole arc of its rise and decline. While I previously showed you the aggregated power index for the British empire, the chart below shows the eight powers that make it up. It shows these from the ascent around 1700 to the decline in the early 1900s. In it, you can see the story behind the rise and decline.
[42]The British Empire and the British PoundBefore we get to the collapse of the British empire and the British pound, let’s take a quick look at the whole arc of its rise and decline. While I previously showed you the aggregated power index for the British empire, the chart below shows the eight powers that make it up. It shows these from the ascent around 1700 to the decline in the early 1900s. In it, you can see the story behind the rise and decline. 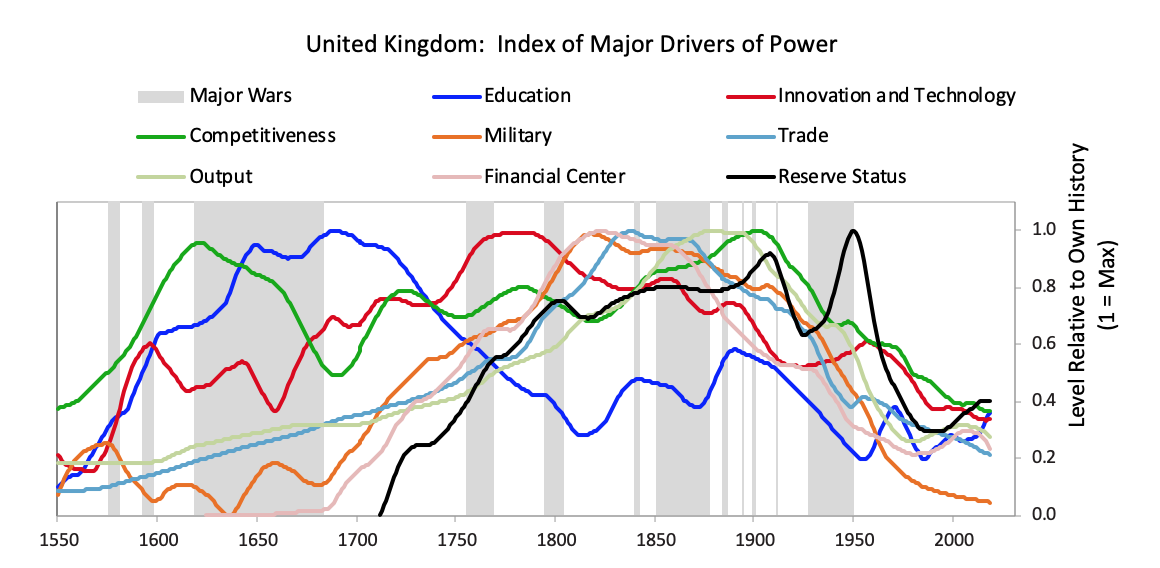 The British empire’s rise began before 1600, with steadily strengthening competitiveness, education, and innovation/technology—the classic leading factors for a power’s rise. As shown and previously described, in the late 1700s the British military power became pre-eminent and it beat its leading economic competitor and the leading reserve currency empire of its day in the Fourth Anglo-Dutch War. It also successfully fought other European rivals like France in a number of conflicts that culminated in the Napoleonic Wars in the early 1800s. Then it became extremely rich by being the dominant economic power. At its peak in the 19th century, the UK’s 2.5% of the world’s population produced 20% of the world’s income, and the UK controlled over 40% of global exports. This economic strength grew in tandem with a strong military, which, along with the privately driven conquests of the British East India Company, drove the creation of a global empire upon which “the sun never set,” controlling over 20% of the world’s land mass and 25% of the global population prior to the outbreak of World War I. With a lag, as is classic, its capital—London—emerged as the global financial center and its currency—the pound—emerged as the leading global reserve currency. As is typical its reserve status remained well after other measures of power started declining in the late 19th century and as powerful rivals like the US and Germany rose. As shown in the chart above, almost all of the British empire’s relative powers began to slip as competitors emerged around 1900. At the same time wealth gaps were large and internal conflicts over wealth were emerging. As you know, despite winning both World War I and World War II the British were left with large debts, a huge empire that was more costly than profitable, numerous rivals that were more competitive, and a population that had big wealth gaps which led to big political gaps. As I previously summarized what happened in the 1914 to post-World War II period, I will skip ahead to the end of World War II in 1945 and the start of the new world order that we are now in. I will be focusing on how the pound lost its reserve currency status. Although the US had overtaken the UK militarily, economically, politically, and financially long before the end of World War II, it took more than 20 years after the war for the British pound to fully lose its status as an international reserve currency. Just like the world’s most widely spoken language becomes so deeply woven into the fabric of international dealings that it is difficult to replace, the same is true of the world’s most widely used reserve currency. In the case of the British pound, other countries’ central banks continued to hold a sizable share of their reserves in pounds through the 1950s, and about half of all international trade was denominated in sterling in 1960. Still, the pound began to lose its status right at the end of the war because smart folks could see the UK’s increased debt load, its low net reserves, and the great contrast with the United States’ financial condition (which emerged from the war as the world’s pre-eminent creditor and with a very strong balance sheet). The decline in the British pound was a chronic affair that happened through several significant devaluations over many years. After efforts at making the pound convertible failed in 1946-47, the pound devalued by 30% against the dollar in 1949. Though this worked in the short term, over the next two decades the declining competitiveness of the British led to repeated balance of payments strains that culminated with central banks actively selling sterling reserves to accumulate dollar reserves following the devaluation of 1967. Around this time the deutschmark began to re-emerge and took the pound’s place as the second-most widely held reserve currency. The charts below paint the picture.
The British empire’s rise began before 1600, with steadily strengthening competitiveness, education, and innovation/technology—the classic leading factors for a power’s rise. As shown and previously described, in the late 1700s the British military power became pre-eminent and it beat its leading economic competitor and the leading reserve currency empire of its day in the Fourth Anglo-Dutch War. It also successfully fought other European rivals like France in a number of conflicts that culminated in the Napoleonic Wars in the early 1800s. Then it became extremely rich by being the dominant economic power. At its peak in the 19th century, the UK’s 2.5% of the world’s population produced 20% of the world’s income, and the UK controlled over 40% of global exports. This economic strength grew in tandem with a strong military, which, along with the privately driven conquests of the British East India Company, drove the creation of a global empire upon which “the sun never set,” controlling over 20% of the world’s land mass and 25% of the global population prior to the outbreak of World War I. With a lag, as is classic, its capital—London—emerged as the global financial center and its currency—the pound—emerged as the leading global reserve currency. As is typical its reserve status remained well after other measures of power started declining in the late 19th century and as powerful rivals like the US and Germany rose. As shown in the chart above, almost all of the British empire’s relative powers began to slip as competitors emerged around 1900. At the same time wealth gaps were large and internal conflicts over wealth were emerging. As you know, despite winning both World War I and World War II the British were left with large debts, a huge empire that was more costly than profitable, numerous rivals that were more competitive, and a population that had big wealth gaps which led to big political gaps. As I previously summarized what happened in the 1914 to post-World War II period, I will skip ahead to the end of World War II in 1945 and the start of the new world order that we are now in. I will be focusing on how the pound lost its reserve currency status. Although the US had overtaken the UK militarily, economically, politically, and financially long before the end of World War II, it took more than 20 years after the war for the British pound to fully lose its status as an international reserve currency. Just like the world’s most widely spoken language becomes so deeply woven into the fabric of international dealings that it is difficult to replace, the same is true of the world’s most widely used reserve currency. In the case of the British pound, other countries’ central banks continued to hold a sizable share of their reserves in pounds through the 1950s, and about half of all international trade was denominated in sterling in 1960. Still, the pound began to lose its status right at the end of the war because smart folks could see the UK’s increased debt load, its low net reserves, and the great contrast with the United States’ financial condition (which emerged from the war as the world’s pre-eminent creditor and with a very strong balance sheet). The decline in the British pound was a chronic affair that happened through several significant devaluations over many years. After efforts at making the pound convertible failed in 1946-47, the pound devalued by 30% against the dollar in 1949. Though this worked in the short term, over the next two decades the declining competitiveness of the British led to repeated balance of payments strains that culminated with central banks actively selling sterling reserves to accumulate dollar reserves following the devaluation of 1967. Around this time the deutschmark began to re-emerge and took the pound’s place as the second-most widely held reserve currency. The charts below paint the picture. 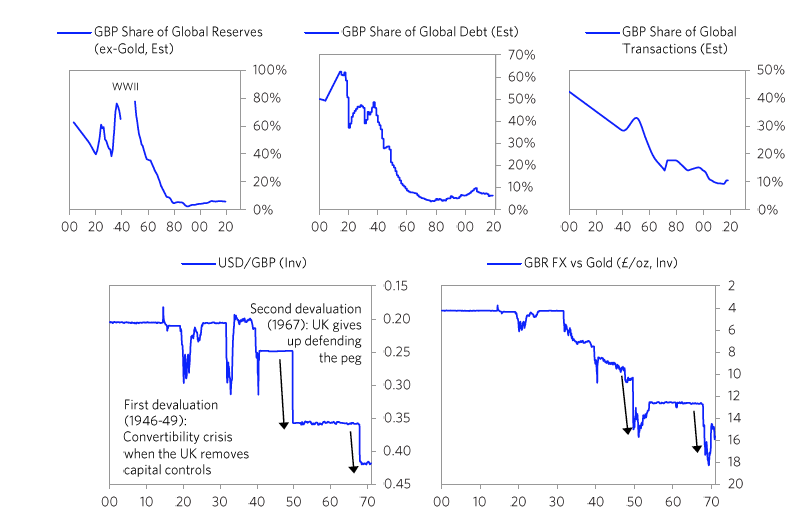 On the following pages we will cover in greater detail the specific stages of this decline, firstly with the convertibility crisis of 1947 and the 1949 devaluation, secondly with the gradual evolution of the pound’s status relative to the dollar through the 1950s and early 1960s, and thirdly with the balance of payments crisis of 1967 and subsequent devaluation. We will focus in on the currency crises. 1) The Pound’s Suspended Convertibility in 1946 and Its Devaluation in 1949The 1940s are frequently referred to as “crisis years”
On the following pages we will cover in greater detail the specific stages of this decline, firstly with the convertibility crisis of 1947 and the 1949 devaluation, secondly with the gradual evolution of the pound’s status relative to the dollar through the 1950s and early 1960s, and thirdly with the balance of payments crisis of 1967 and subsequent devaluation. We will focus in on the currency crises. 1) The Pound’s Suspended Convertibility in 1946 and Its Devaluation in 1949The 1940s are frequently referred to as “crisis years”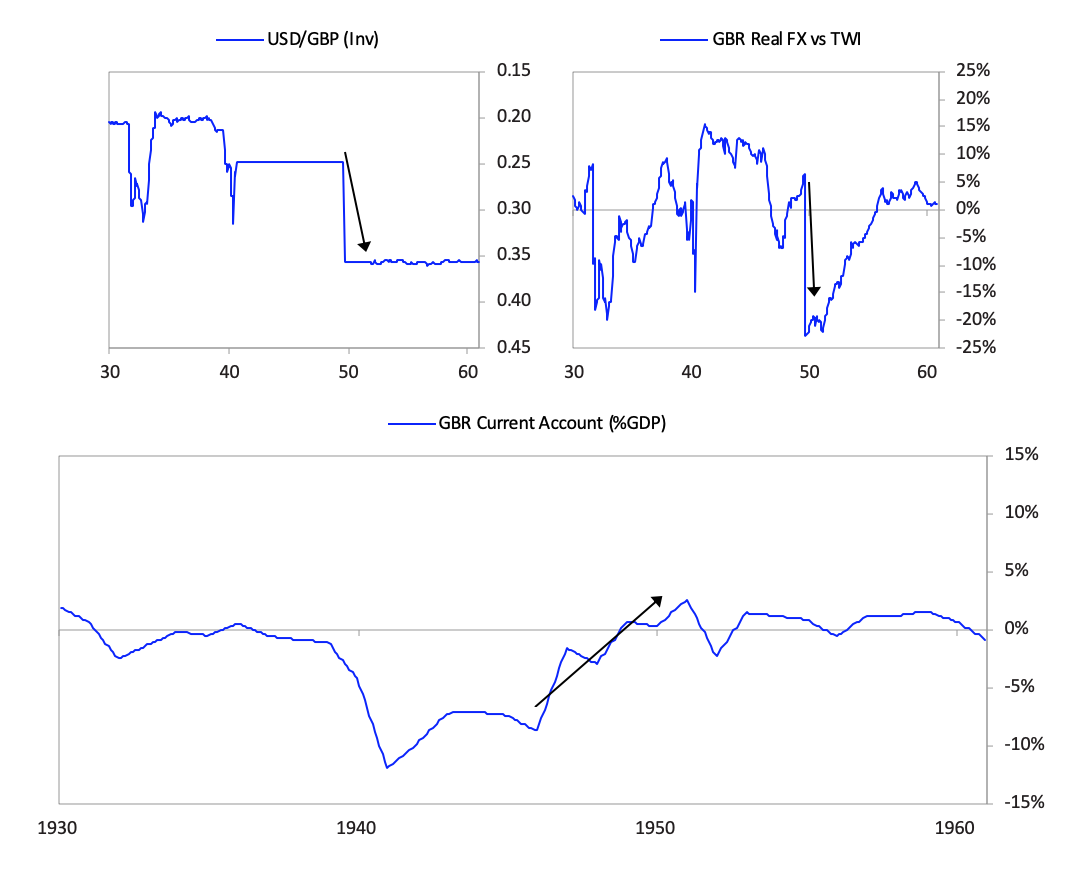 The currency move, which devalued sterling debt, did not lead to a panic out of sterling debt as much as one might have expected especially in light of how bad the fundamentals for sterling debt remained. That is because a very large share of UK assets was held by the US government, which was willing to take the valuation hit in order to restore convertibility, and by Sterling Area economies, such as India and Australia, whose currencies were pegged to the pound for political reasons.
The currency move, which devalued sterling debt, did not lead to a panic out of sterling debt as much as one might have expected especially in light of how bad the fundamentals for sterling debt remained. That is because a very large share of UK assets was held by the US government, which was willing to take the valuation hit in order to restore convertibility, and by Sterling Area economies, such as India and Australia, whose currencies were pegged to the pound for political reasons.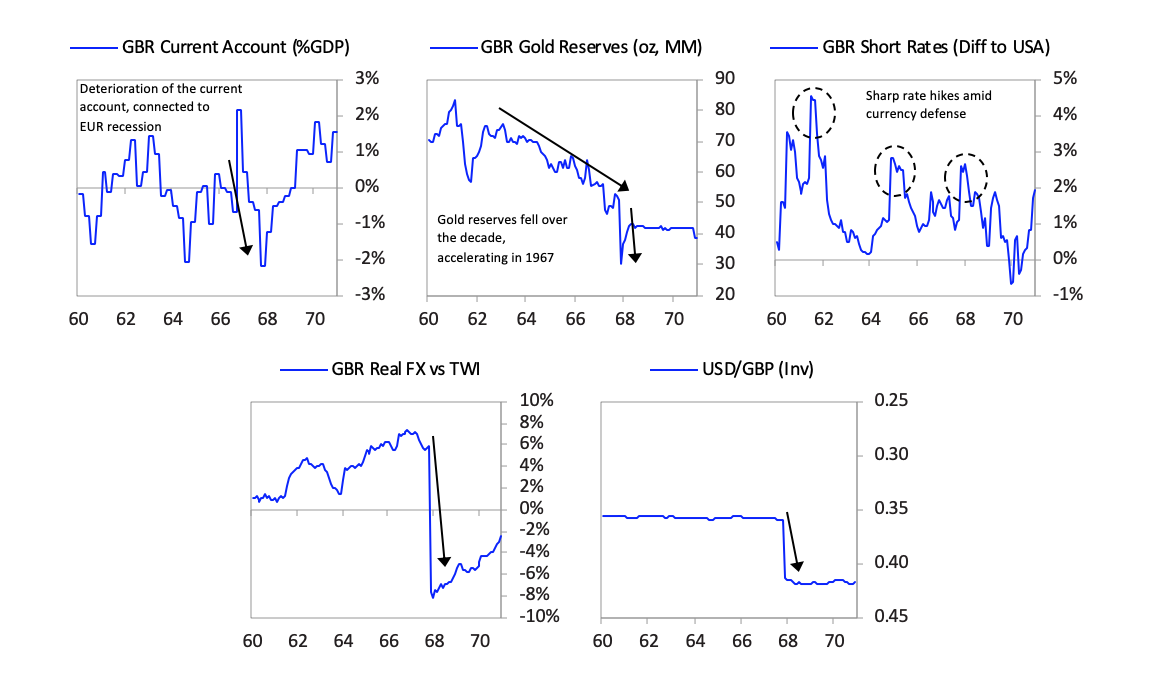 After the devaluation little faith remained in the pound as the second-best reserve currency after the dollar. For the first time since the end of World War II, international central banks began actively selling their sterling reserves (as opposed to simply accumulating fewer pounds in new reserve holdings) and instead began buying dollars, deutschmarks, and yen. As you can see in the chart below on the left, the average share of sterling in central bank reserve holdings collapsed within two years of the devaluation. At the same time the UK was still able to convince Sterling Area countries not to diversify away from the pound. In the Sterling Agreement of 1968, Sterling Area members agreed to maintain a floor on their pound reserve holdings, as long as 90% of the dollar value of these holdings was guaranteed by the British government. So although the share of pound reserves in these Sterling Agreement countries like Australia and New Zealand remained high, this was only because these reserves had their value guaranteed by the British in dollars. So all countries that continued to hold a high share of their reserves in pounds after 1968 were holding de facto dollars with the British bearing the risk of a further sterling devaluation.
After the devaluation little faith remained in the pound as the second-best reserve currency after the dollar. For the first time since the end of World War II, international central banks began actively selling their sterling reserves (as opposed to simply accumulating fewer pounds in new reserve holdings) and instead began buying dollars, deutschmarks, and yen. As you can see in the chart below on the left, the average share of sterling in central bank reserve holdings collapsed within two years of the devaluation. At the same time the UK was still able to convince Sterling Area countries not to diversify away from the pound. In the Sterling Agreement of 1968, Sterling Area members agreed to maintain a floor on their pound reserve holdings, as long as 90% of the dollar value of these holdings was guaranteed by the British government. So although the share of pound reserves in these Sterling Agreement countries like Australia and New Zealand remained high, this was only because these reserves had their value guaranteed by the British in dollars. So all countries that continued to hold a high share of their reserves in pounds after 1968 were holding de facto dollars with the British bearing the risk of a further sterling devaluation.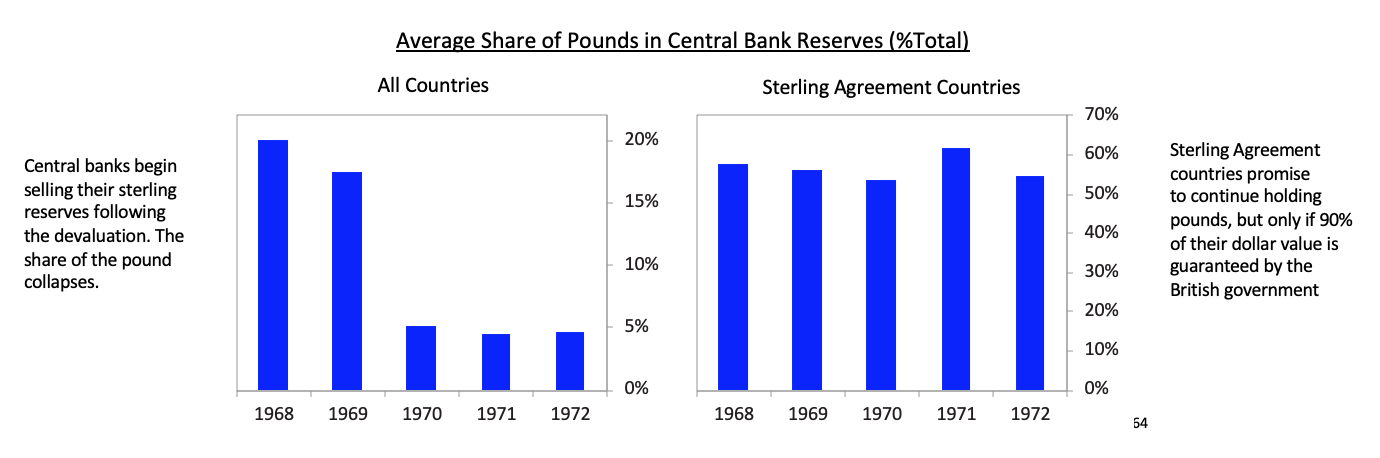 [64]By this time the dollar was having its own set of balance of payments and currency problems, but that is for the next installment of this series when I turn to the United States and China.
[64]By this time the dollar was having its own set of balance of payments and currency problems, but that is for the next installment of this series when I turn to the United States and China. {kind=link}